

年齡越大,死亡風險越高。自然界似乎都跟隨著這個常態,這被稱為岡珀茨-梅卡姆死亡率定律(Gompertz–Makeham law of mortality)。但是在這個世界上,有一種近乎完美的生物不被這個規則所束縛。牠就是裸鼴鼠(Heterocephalus glaber)。
裸鼴鼠是一種分布在東非地區的挖掘類齧齒目動物,也是目前被分類於裸鼴鼠屬下的唯一物種。裸鼴鼠是一種真社會性哺乳動物另外一種是達馬拉蘭鼴鼠)。
有一隻巨大的鼠后以及少數雄鼠。其餘全是工鼠,工鼠會因為鼠后給予的排泄物中的激素生殖能力受到抑制。牠們負責採集食物,為鼠后保暖,挖掘隧道,抵禦外敵等功能。
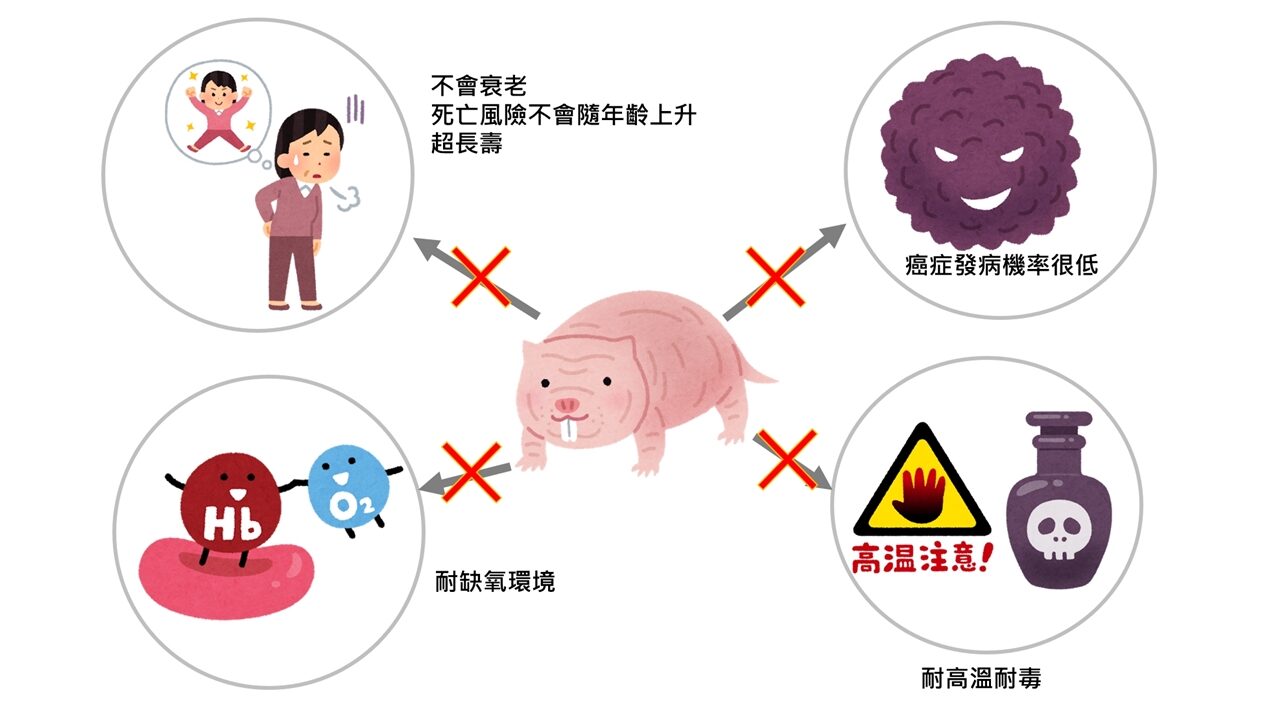
圖1. 接近完美的生物-裸鼴鼠
有關裸鼴鼠的生態可以參照天地創造設計部動畫第10集。
但是這種動物會受到關注的最大特徵在於:
隨著年齡增長死亡率遠低於其他動物。
癌症發病機率極低。
身體機能幾乎不會退化。
到死都能繁殖後代。
不怕有毒植物。
不須氧氣也能活存一段時間。
哺乳類動物中唯一可以調節體溫的物種,能耐高溫。
極低的代謝率,壽命是一般老鼠的10倍,約30年壽命。
無皮膚痛覺。
與人類DNA相似度為80-93%。
也就是說,裸鼴鼠是人類追求抗老化機制的最佳模式生物,牠們犧牲了基因的變異性誕生了極為長壽健康的生理機制,因此是哺乳類中最接近人類永生願望的最佳研究對象。
裸鼴鼠與人類
在2019年的Phenoptosis期刊中,N. A. Popov & V. P. Skulachev共同發表了一篇研究報導整理出了裸鼴鼠與人類的異同之處。
| 組織層次 | 裸鼴鼠 | 人類 | 人和裸鼴鼠的共同特徵 |
| 1 | 2 | 3 | 4 |
| 生物特徵 | 體重比濱鼠科的 20 種相關物種小得多。 沒有毛髮(在成年囓齒動物中是獨一無二的)。 對於體重相當的動物,妊娠期比預期的要長得多。 比其他囓齒動物長得多的生長和成熟時間。 隨著年齡的增長,生殖能力增加(與在其他哺乳動物中觀察到的減少相比)。 維持恆定體溫的能力有限(類似於新生動物)。 與年齡相關的疾病,如癌症、糖尿病、心血管疾病、肝臟和大腦病變、腎臟疾病和許多感染,作為裸鼴鼠死亡的原因非常罕見或不存在(就像哺乳動物幼崽的情況一樣)。 骨骼系統的延遲成熟。 高最大壽命; 死亡率低(與其他囓齒動物相比),至少在生命的前 24 年中僅增加微不足道。 沒有與年齡相關的認知功能下降。 | “平面”,正頜。 體毛減少或缺失。 枕骨大孔的中心位置(在大多數其他靈長類動物的個體發育過程中向後遷移)。 保留顱骨結構直到高齡。 手和腿的結構。 骨盆的形狀。 女性陰道在腹側位置。 牙齒排列和頭顱縫隙的某些變化。 沒有顱嵴。 沒有眉脊。 顱骨薄。 顱腔下眼眶的位置。 短頭畸形。 小牙齒。 牙齒萌生晚。 拇指不可能完全旋轉。 長期嬰兒依賴。 長期的成長。 壽命長。 與體型相比四肢短。 腿比手臂長。 身體的垂直位置。 與黑猩猩相比,妊娠時間更長。 | 毛髮缺失或減少。 長壽命和健康壽命。 長期的成長。 妊娠期長。 長期無助的嬰兒期。 性成熟期長。 |
表1. 生物特徵比較。
| 1 | 2 | 3 | 4 |
| 組織 | 沒有耳廓,與其他成年哺乳動物相比就像新生兒一樣。 沒有陰囊,如新生兒,海馬 CA3 區鈣結合蛋白如新生兒型分佈。 海馬對外源性腺苷的敏感性低。 與其他囓齒動物相比,大腦發育的時間很長。 肺的許多形態學方面發育不全。 新生裸鼴鼠大腦是新生小鼠大腦的兩倍大,新生小鼠大腦在出生時約為成年體重的 17%,但到 2 週時達到成年體重的 90%,而裸鼴鼠大腦在出生時約為成年體重的 41%,但直到達到成人體重的 90% 3個月大。在斷奶期間,裸鼴鼠的大腦明顯大於小鼠,但成年裸鼴鼠的大腦在考慮體重時明顯小於小鼠。 裸鼴鼠出生時具有明確定義的齒狀回 (DG) 分子層,其中表達 Sox2 的細胞明顯較少,而新生小鼠大腦中幾乎所有齒狀回細胞都是 Sox2 陽性的。這兩個物種出生時在心室和心室下區(分別為 VZ 和 SVZ)中表達 Sox2 的細胞水平相當,但在出生後的第一個星期,該水平在小鼠中急劇下降,但在裸鼴鼠中,該水平在出生後與3歲時沒有顯著變化。 腦神經元在復氧期間對缺氧和隨後的氧化應激具有非常高的抵抗力(類似於新生動物)。 與其他成年哺乳動物的影響相比,缺乏突觸配對脈衝刺激。 保留成年期神經元再生和伸長的能力。 肺神經上皮體,通常在出生後第一周數量減少,在超過兩週齡的裸鼴鼠中大量出現。 血管彈性不會隨著年齡的增長而降低。 隨著年齡的增長,大腦皮層區域沒有減少。 沒有與年齡相關的骨骼礦物質密度降低。 隨著年齡的增長,關節軟骨的狀態沒有任何下降。 與在其他囓齒動物中觀察到的變化相反,裸鼴鼠平滑肌對 NO 的敏感性不會隨著年齡的增長而降低。 在小鼠中,皮質層 1 直到 2 週齡才變得清晰,並且在 3 週時失去 DCX 表達。相比之下,裸鼴鼠出生時皮層排列更有序,皮層發育更發達,但皮層成熟期比小鼠長。 小鼠的神經放電模式水平在出生後的 6 個月內達到最大值,而在裸鼴鼠中,這一水平在第一個生命的十年中緩慢增加。 | 耳廓的形狀。 內眥褶。 腦量比較大。 女性生殖器陰唇。 沒有棒(陰莖骨)。 小鼻子。 人類保留了更原始的骨骼和肌肉形狀,而在猴子中,這些器官的解剖結構更加複雜和專業。 人類的大腦比黑猩猩的大腦大得多。 人類大腦的生長速度在出生前一直在增加,而在黑猩猩中,這個參數開始下降得更早。 在成年人中,神經元保留了明確的“次要”特徵,例如增加的突觸活動和可塑性,以及生命最初 20 年的髓鞘化不完全。 與類人猿相比,人類的紡錘形神經元更大,數量也更多。 | 非常不發達的犁鼻器官(類似於新生兒)。 增加突觸活動和可塑性。 出生後幾年中性纖維髓鞘化不完全。 在大腦發育的早期階段突觸的爆炸性形成(“剩餘突觸發生”)。 |
表2. 組織特徵比較。
| 1 | 2 | 3 | 4 |
| 細胞和分子 | 神經元長期興奮時沒有 Ca2+ 過載。 骨骼肌在出生後的統一線粒體系統的發育嚴重延遲。 隨著年齡的增長,線粒體數量及其活性增加(而不是減少)。 與小鼠相比,裸體雄性大鼠不表現出與年齡相關的線粒體呼吸控制下降。 沒有與年齡相關的代謝普遍下降。 在其他衰老哺乳動物中觀察到的脂質過氧化指數和脂質氧化損傷沒有與年齡相關的增加。 裸鼴鼠的 ROS 生成不隨年齡增加(與其他哺乳動物不同)。 裸鼴鼠的軸突成分與人類更相似,裸鼴鼠即使在大腦發育完成後仍保持 3R tau 的表達,小鼠會突然的停止。 產後第 8 天 3R tau 的下調。 在裸鼴鼠中,線粒體的輕度去極化會保留到 10 歲,而在小鼠中,這種線粒體抗 ROS 防禦機制在出生後一年內就停止發揮作用。 裸鼴鼠肝臟線粒體的輕度去極化至少保留到 10 歲,而在小鼠中,它在出生後立即減少。 編碼 FAS 激活的絲氨酸/蘇氨酸激酶 (FASTK) 的基因失活,FASTK 是一種抗細胞凋亡和促炎酶。 超氧化物歧化酶 1 和 2 以及過氧化氫酶的水平沒有與年齡相關的降低。 添加ADP後,成年裸鼠線粒體中腺嘌呤核苷酸水平降低,呼吸水平升高(類似於大鼠和小鼠胚胎和新生兒)。 成年動物中異常高的蛋白酶體活性。 胰島素 B 鏈序列的變化導致激素活性降低到胚胎水平。 成年裸鼴鼠的 IGF2 水平沒有降低。 成年裸鼴鼠IGF-1、胰島素誘導基因2(INSIG2)表達降低,IGF1R和抵抗素(RETN)基因表達增加。 由於缺乏 P 物質,不會感覺到辣椒素或酸引起的疼痛。 成體中存在谷氨酸 NMDA 受體 GluN2D 亞基,而其他哺乳動物僅在新生兒中存在。 老年動物的葡萄糖耐量沒有降低。 糖化血紅蛋白水平沒有與年齡相關的增加。 細胞間隙中存在高分子量透明質酸。 在裸鼴鼠中,輕鏈神經絲 (NF-L) 的水平在出生後第 2-4 週增加了 33%;到 4 個月時,比新生兒高 168%。 在小鼠中,NF-L 水平在出生後迅速增加,並在出生後第一個星期達到成年水平。 DCX(微管相關蛋白,doublecortin)在小鼠中的表達高於裸鼴鼠,但在齒狀回和腦室下區均迅速降低,而在裸鼴鼠中,其表達量逐漸降低。 Map2D 亞型(微管相關蛋白)的水平在裸鼴鼠中保持穩定,直到 6 個月大,而在大鼠中,這種蛋白的表達在出生後弱 3 時完全下降。 裸鼴鼠的突觸素水平在 2-4 個月至 9 個月大時顯著增加,而在小鼠中,這發生在出生後的第 10-35 天。 裸鼴鼠大腦中酪氨酸羥化酶的表達從出生到 3 歲逐漸增加,而在大鼠中,這種情況發生得更快 – 在發育的第 4-5 週。 與小鼠不同,裸鼴鼠在出生時表達髓鞘相關糖蛋白 (MAG)。 在小鼠中,直到 3 個月大時,己糖激酶 II 在肌肉線粒體中的活性高於更活躍的己糖激酶 I。 在裸鼴鼠中直到 10 年後才發生。 | 這兩個物種在 p53 富含脯氨酸的域中具有最高數量的 PXXP 基序(人類 5 個,裸鼴鼠 4 個)。 酪氨酸羥化酶的水平會增加,直到 2-3 歲。 3R tau 蛋白的穩定表達。 |
表3. 細胞與分子表現比較。
結論:從期刊整理過去研究可以得出,裸鼴鼠與人類類似成長期時間很長。許多從新生兒到成年會逐漸衰退的基因或粒線體表現並沒有下降。對於活性氧類(Reactive oxygen species,ROS)的生成也不隨年齡上升,減少了DNA損壞的機率。這可能給予我們後續的研究方向,也就是延長某些只在新生兒時期表現的基因,提升成年人抗ROS的能力。
Neotenic Traits in Heterocephalus glaber and Homo sapiens
參考文獻
Popov, N. A., & Skulachev, V. P. (2019). Neotenic traits in Heterocephalus glaber and Homo sapiens. Biochemistry (Moscow), 84(12), 1484-1489.





